Welcome to the forums at seaphages.org. Please feel free to ask any questions related to the SEA-PHAGES program. Any logged-in user may post new topics and reply to existing topics. If you'd like to see a new forum created, please contact us using our form or email us at info@seaphages.org.
Recent Activity
All posts created by delesall
| Link to this post | posted 11 Jan, 2018 18:33 | |
|---|---|

|
Hi all I just downloaded DNAMaster from the Baylor site (https://baylor.app.box.com/v/DNAMasterMac) using the version from 2017.5.23 I have been able to upload a sequence but when I do an auto-annotation (no blasting) nothing appears in the features? Any advice? And has somebody compiled a document on issues and things to know about running DNAMaster on a mac and is willing to share? Thanks and happy annotating in 2018! Veronique |
Posted in: DNA Master → Running DNA Master on a Mac using Wine
| Link to this post | posted 11 Aug, 2017 21:27 | |
|---|---|
|
|
So, I should have better phrased my question. Do we want the prob of a CCM to be greater than 0 or greater than as certain value? If so, what is that threshold? prob of 0.2 or 0.5? 
|
Posted in: Functional Annotation → cluster J terminases
| Link to this post | posted 11 Aug, 2017 03:34 | |
|---|---|
|
|
The Lysin A in Gordonia phage is encoded by two genes, representing two domains. In Gordonia phage Adgers, these two genes correspond to N-acetyl-BetaD-muramidase domain (pfam 01510) and to a peptidase domain (pfam 01551). In Gordonia phage SuperSulley, the two Lysin A genes are ORF 49 (start 37820) and ORF 50 (start 38485) and respectively correspond to a peptidase domain (pfam 13529) and to a glycoside hydrolase (cd00325). See Payne and Hatfull (2012 PlosOne - Mycobactteriophage Endolysins…  . pfam 13529 is not listed in Table 1! . pfam 13529 is not listed in Table 1!
|
Posted in: Functional Annotation → Lysins in Gordonia
| Link to this post | posted 11 Aug, 2017 01:54 | |
|---|---|
|
|
In reviewing the annotation of phage LilHazelnut, I noticed two genes, ORF 68, start 45763, and ORF 69, start 46743, with decent HHPred matches (98% prob, 40% coverage, reasonably low E-values) to respectively G38P, the N-terminal phage replisome organiser (The protein contains a region of low-complexity sequence that reflects DNA direct repeats able to function as an origin of phage replication) and G39P, Loader and inhibitor of phage G40P. G39P inhibits the initiation of DNA replication by blocking G40P replicative helicase. Although the protein of ORF 70 does not have matches to a replicative helicase, should we call some function for ORF 68 and 69, and if so, what? From Ayora et al (1999) J Mol Biol. 288(1):71-85) Abstract Initiation of Bacillus subtilis bacteriophage SPP1 replication requires the phage-encoded genes 38, 39 and 40 products (G38P, G39P and G40P). G39P, which does not bind DNA, interacts with the replisome organiser, G38P, in the absence of ATP and with the ATP-activated hexameric replication fork helicase, G40P. G38P, which specifically interacts with the phage replication origin (oriL) DNA, does not seem to form a stable complex with G40P in solution. G39P when complexed with G40P-ATP inactivates the single-stranded DNA binding, ATPase and unwinding activities of G40P, and such effects are reversed by increasing amounts of G38P. Unwinding of a forked substrate by G40P-ATP is increased about tenfold by the addition of G38P and G39P to the reaction mixture. The specific protein-protein interactions between oriL-bound G38P and the G39P-G40P-ATPgammaS complex are necessary for helicase delivery to the SPP1 replication origin. Formation of G38P-G39P heterodimers releases G40P-ATPgammaS from the unstable oriL-G38P-G39P-G40P-ATPgammaS intermediate. G40P-ATPgammaS binds to the origin region, the uncomplexed G38P fraction remains bound to oriL, and the G38P-G39P heterodimer is lost from the complex. We demonstrate that G39P is a component of an oligomeric nucleoprotein complex which plays an important role in the initiation of SPP1 replication. |
Posted in: Functional Annotation → Replisome organizer/
| Link to this post | posted 10 Aug, 2017 19:29 | |
|---|---|
|
|
In reviewing the annotation of Demsculpinboyz (an F2),the protein of ORF 82 (start 43366, stop 44214) has blast matches to tRNA methyl transferase, PAPs reductase, and ribonucleotide reductase! There is no CDD hits but a number of good HHPred matches (Prob 99%, coverage greater than 65%, low E-values)to 1) phosphoadenosine phosphosulfate sulfurtransferase, 2) Adenine nucleotide alpha hydrolases superfamily, and 3) adenosine 5'-phosphosulfate reductase. All this suggests that going with ribonucleotide reductase is safest bet. What does the forum think? |
Posted in: Functional Annotation → Methyl transferase, Reductase or ?
| Link to this post | posted 10 Aug, 2017 18:19 | |
|---|---|
|
|
For the output, are we just looking for the absence or presence of a coiled coil motif? |
Posted in: Functional Annotation → cluster J terminases
| Link to this post | posted 03 Jul, 2017 17:30 | |
|---|---|
|
|
I would still want to call both genes - the "original" gene in other phages as no known function. The question is whether I annotate both as "hypothetical protein" or "hypothetical protein, N-terminus" and "hypothetical protein, C-terminus". And what Genbank will accept… |
Posted in: Gene or not a Gene → Gene split in 2
| Link to this post | posted 16 Jun, 2017 20:30 | |
|---|---|
|
|
Thanks Chris That does look like a possible answer to the problem. I think we may need to go back to BYU for the sequencing data! |
Posted in: Gene or not a Gene → Gene split in 2
| Link to this post | posted 06 Jun, 2017 15:50 | |
|---|---|
|
|
Yes - see attached file - 34B is (obviously) between labeled 34 and 35 |
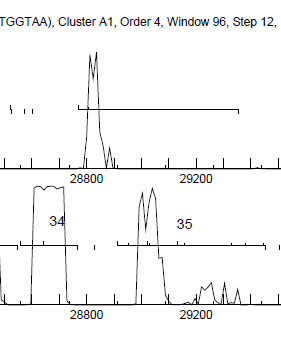 9Kb
9KbPosted in: Gene or not a Gene → Gene split in 2
| Link to this post | posted 06 Jun, 2017 15:38 | |
|---|---|
|
|
From HHPred (as well as Interpro, pfam, … Ead/Ea22-like protein This family contains phage proteins and bacterial proteins that are likely to represent integrated phage proteins. This family includes the Lambda phage Ea22 early protein as well as the Bacteriophage P22 Ead protein. A quick Google search does not find more information than this… I'm happy to just say NKF |
Posted in: Functional Annotation → Ead/Ea22-like protein