Welcome to the forums at seaphages.org. Please feel free to ask any questions related to the SEA-PHAGES program. Any logged-in user may post new topics and reply to existing topics. If you'd like to see a new forum created, please contact us using our form or email us at info@seaphages.org.
Recent Activity
All posts created by anders
| Link to this post | posted 21 Jul, 2024 21:35 | |
|---|---|

|
Genes related to MidnightRain_54 are often called by Glimmer and GeneMark to start near the beginning of strong coding potential, leaving a sizeable gap. But the reading frame is open upstream and can be extended to reduce the gap to ~16 bp to the upstream gene. These upstream starts are conserved across the pham (as of June 2024). When the upstream starts are chosen, the longer proteins can have hits on HHPred to metallopeptidases (like IrrE) and to helix-turn-helix domain containing proteins. The location of the called start can influence the alignment. Shorter proteins often match the HTH domain well, but only part of the peptidase domain. Longer proteins match the full peptidase domain, which includes part that is HTH-like. Try HHPred on proteins of different lengths. If the longer proteins continue to add alignment to metallopeptidases, the longer protein may be the better call. See the attachment to visualize the alignments. |
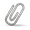 773Kb
773Kb| Link to this post | posted 21 Jul, 2024 21:13 | |
|---|---|
|
|
A gene at the far right end may not have much coding potential across its whole ORF, and it might not be called by Glimmer or GeneMark. It is worth looking to see if there is an open reading frame that encodes an HNH endonuclease. And worth checking to see if the ORF is longer than the coding potential would suggest. Attachment shows some example coding potential plots from GeneMark. |
Posted in: Cluster AY Annotation Tips → Right-most gene may be an HNH
| Link to this post | posted 21 Jul, 2024 21:07 | |
|---|---|
|
|
A reverse gene is often sandwiched between tail assembly chaperone (TAC) genes and the tape measure gene. Look for space and good coding potential. Some AYs don't have room for a gene between the TACs and tapemeasure, such as AbbyDaisy and Raqqa. |
| Link to this post | posted 07 Nov, 2023 06:25 | |
|---|---|
|
|
In the other thread about immunty repressors (https://seaphages.org/forums/topic/4952/), Debbie invited folks to post wet bench data. My students and I have some preliminary evidence that the A1-like immunity repressor in the F1 phages Coco12 and Phanphagia is sufficient to defend against the A1 phage Bxb1. Briefly, –they made lysogens of five F1 phages, and found that lysogens of Coco12 and Phanphagia reduced plaque formation of superinfecting A1 phage Bxb1 (EOP ~10^-3). The other three lysogens (carrying GUmbie, Gandalph, and Veteran) had no effect on Bxb1 superinfection. –They observed that Coco12 and Phanphagia carry the A1-like immunity repressor gene, wereas the other F1 phages in their tests do not. Perhaps this A1 repressor gene is causing the defense against Bxb1. –They cloned the A1-like repressor ORF into the expression plasmid pLAM12, and they cloned the two gene operon including the A1-like repressor and its upstream gene into the integration plasmid pMH94. They transformed M.smeg with the plasmids and tested for Bxb1 plaque formation. –Both plasmid clones appear to be sufficient for defense against Bxb1 infection, compared to strains carrying empty plasmids. I'm attaching a summary of the experiments. |
| Link to this post | posted 09 Jun, 2023 22:08 | |
|---|---|
|
|
As I tried to apply this to some proteins that we are annotating, I was confused because I couldn't replicate Rick's result with Q1 being in the first alpha helix.Pollenz When I used the protein sequence that Tejas posted, I got HHPred alignments that did predict some beta sheet in front of the first alpha helix. Then I realized that Rick may have used the draft genome protein that began at the Glimmer auto-annotated start, which was downstream of the start that Tejas had used for the amino acid sequence she posted. If we use a start codon even further upstream, to make the protein longer at its N-terminus, we can get a pretty good HHPred alignment to ribbon-helix-helix structures that includes a full-length beta strand. I've attached an image of the HHPred alignment with the longer amino acid sequence. My interpretation is that HHPred alignments do support the ribbon-helix-helix domain, if translation begins at a start codon far enough upstream to contain the predicted beta strand. For Usavi, it will come down to which start codon the annotators choose as the most appropriate start. For the genes we're working on, we will pay close attention to the key features of ribbon-helix-helix that Rick nicely explained above! ~Kirk |
| Link to this post | posted 20 May, 2021 19:41 | |
|---|---|
|
|
This is a follow-up to the toxin BrnT posts https://seaphages.org/forums/topic/4458/. Like Paola (K5), SoSeph (K5) also has a pair of genes that looks like the BrnTA toxin/antitoxin pair of genes [SoSeph_draft_88 (57177-57455) and SoSeph_draft_89 (57457-57888 )]. As Jordan, Veronique and Kristen described in the BrnT discussion, the downstream antitoxin gene has a ribbon-helix-helix structure. In HHPred, it hits the KacA antitoxin crystal in and RelD and CopG antitoxin CCD domains, all of which are also members of type II toxin-antitoxin systems. We propose that this ribbon-helix-helix protein be called "antitoxin in toxin/antitoxin system, BrnA-like". It has the predicted ribbon-helix-helix motif of the antitoxin, the predicted size (ie, larger than the toxin), and the predicted synteny (immediately downstream of the toxin in a potential two-gene operon). BrnTA structure-function paper (Heaton et al., 2012) https://www.jbc.org/article/S0021-9258(20)53133-5/fulltext The official function list includes the BrnT toxin but does not include the partner BrnA antitoxin. |
Posted in: Request a new function on the SEA-PHAGES official list → antitoxin in toxin/antitoxin system, BrnA-like
| Link to this post | posted 14 May, 2021 19:14 | |
|---|---|
|
|
Thanks for this, Chris. I would like to change my suggestion to the 34328 start! Knowing that mass spec data exist, I've been applying the "second-of-tandem-start-codons" rule when there is a tandem set at 4bp and 1bp overlaps, choosing the 1 bp overlap start. In my mind, I had thought the data applied to cases where a ribosome was potentially restarting after termination of the upstream gene (4 bp and 1 bp overlaps). But the statement you quoted in the guide suggests it is more broadly observed. Which makes me wonder: are the mass spec data relative to tandem start codons published someplace? It would be interesting to look at the range of cases observed. |
Posted in: Choosing Start Sites → F1 gene needs help on start site
| Link to this post | posted 13 May, 2021 22:26 | |
|---|---|
|
|
Hi, here's how I think about this. I see that the coding potential extends further upstream than the Glimmer start at 34172, which strongly suggests that the start is further upstream. So the only possible starts are two that are right next to each other upstream of this, 34331 and 34328. Neither of them have a significant match to a Shine-Dalgarno sequence, so they seem equal with regard to SD. To me, it seems like an arbitrary choice to choose one over the other, and there would be a very minor difference in the protein of one amino acid at the N-terminus. Sounds like GM called the 34331 site in your analysis (it did not in my DNA Master auto-annotation just now). This makes the longest possible ORF and would match 1:1 with the start of SiSi_42 and two others. This would tip the balance for me and I would choose the 34331 start. Best wishes! -Kirk |
Posted in: Choosing Start Sites → F1 gene needs help on start site
| Link to this post | posted 07 Jan, 2021 23:38 | |
|---|---|
|
|
Hi Maria, The official function list does have Holliday junction resolvase. I found it near the bottom of the spreadsheet when we annotated some M2s recently. Best, Kirk |
| Link to this post | posted 09 Dec, 2020 00:02 | |
|---|---|
|
|
Hi, We're curious to hear what others think about this situation. We're working on CrunchyBoi and PineapplePluto and here's what we see. CrunchyBoi, PineapplePluto, and Pabst (EK1) share a ~100 bp insertion on the right end of the genome relative to other EKs. Glimmer calls a gene in this region in all three, and there is coding potential across the orf in self-trained Genemark. The orf fills the gap created by the insertion. Its overlaps are a bit large (7 bp overlap with the upstream gene, and 14 bp overlap with the downstream gene). SD z=2.5. Nucleotide substitutions in the ORF among the three genomes change 14 codons: 10/14 codon changes are synonymous and do not alter the AA, and 3 of the 4 nonsynonymous changes make chemically conserved AA changes (V->A, N->D, E->K; non-conserved = W->G). We might expect to see this pattern if natural selection is keeping this ORF open and producing a protein. (I have attached views of the data.) If the orf was in the middle of a genome, we would probably call it as a small gene. But the EKs are circularly permuted, and this reverse ORF sits over the boundary at the genome ends, causing us to proceed with more caution. Should we call it a gene? thanks! Kirk |
Posted in: Gene or not a Gene → gene at end of some EK1s?